Startseite
Einführung
Erdzeitalter
Fossilisation
Morphologie
Rekonstruktion
Sehapparat
Einrollung
Häutung
Bestachelung
Wachstum
Ernährung
Reproduktion
Pathologie
Ordnungen
Ursprünge
Aussterben
Fundorte
Kontinentaldrift
Präparation
Galerien
Gastsammlung
Fälschungen!
Literatur
Trilo-Links
Evolution
Kreationismus
Glossar
Impressum
Über den Autor

![]()

Nahrungsbeschaffung Was gab es bei "Trilobitens" zum Frühstück?
• Spurenfossilien, sog. "trace fossils" wie z. B. Rusophycus und Cruziana, bei denen es sich nicht um die eigentlichen Überreste von Trilobiten handelt, sondern um Hinterlassenschaften in Form von Bewegungsspuren im Sediment aus denen man auf ein bestimmtes Verhaltensmuster schließen kann.
Fortey und Owens haben bereits 1999 in ihrem Aufsatz (Feeding habits in trilobites, Palaeontology, Vol. 42, Part 3, pp. 429-465) die Auffassung vertreten, daß sich aus einer Kombination verschiedener Charakteristika aus der oben angeführten Aufzählung ein Bild ergeben kann. Sie teilten demnach die Trilobiten in verschiedene Gruppen auf, was die Nahrungsaufnahme betraf: • Beutegreifer und Aasfresser
Insbesondere dem Hypostom kommt nach ihrer Auffassung Bedeutung zu. Sie geben zu Bedenken, daß das Hypostom auch als Teil des Mundapparates anzusehen ist und dabei half, Nahrung zuzuführen. Rezente wie auch archaische, fossiliert vorliegende Beutegreifer und Aasfresser besitzen in der Regel Mundwerkzeuge, die es ihnen ermöglichen, Beute festzuhalten, aufzubrechen, zu zerkleinern, etc. Dies erfordert eine gewisse Stabilität und Ausformung. Nun gibt es ja verschiedene Arten von Hypostomen. Konterminante und impendente Hypostome sind mittels Naht dennoch fest mit der Rostralplatte, in seltenen Fällen direkt mit der vorderen Cephalondublüre verbunden, die regelmäßig stark ausgebildet ist (z. B. Asaphus), d. h. sie besitzen eine belastbare Basis. Oft weisen diese Hypostome zusätzlich eine Vielzahl an unterschiedlichen Ausformungen von Gabeln, Zähnen, oder in Einzelfällen sogar raspelartige Auswüchsen, etc., auf. Hinzu kommt, daß die meisten Trilobiten mit derartigen Charakteristika in der Regel zu Fortey und Owens betrachten die Beutegreifer und Aasfresser als Anwender einer primitiven, ursprünglichen Art der Nahrungsaufnahme, die sich allerdings bis weit über das Ordovizium hinaus erstreckte und sowohl in den Paradoxidoidea, Corynexochida, einigen Asaphida, Lichida, Odontopleuridae, Calymenoidea, Phacopina und sekundär-konterminanten Proetida zu beobachten sei. Spurenfossilien, namentlich die bereits erwähnten Rusophycus, die in der Regel größeren Asaphiden zugeordnet werden und deutliche Abdrücke der Hypostomgabeln und Gnathobasen zeigen, treffen des öfteren mit anderen Spuren zusammen, die von wurmartigen Lebewesen zu stammen scheinen, und die mit dem Zusammentreffen enden. Dies deutet ebenfalls darauf hin, daß der Trilobit räuberisch tätig war, auch wenn diese Interpretation in jüngerer Zeit bisweilen angezweifelt worden ist (Rydell et al., 2001: Trace fossil associations in the Swedish Mickwitzia sandstone: Did trilobites really hunt for worms? GFF, Vol. 123 pp. 247-250)
Nach Fortey und Owens handelt es sich bei Trilobiten mit natantem Hypostom also in der Regel um Partikel- oder Suspensionsfresser, die verwertbare organische Reste entweder vom Sediment abweideten oder aus ihm herausfiltrierten, wobei ihnen der "Hypostom-Aufzug" behilflich war. Ihr in der Regel großflächiges Auftreten in Kombination mit ihrer relativ geringen Größe weist unter Beachtung anderer Beobachtungen ebenfalls auf eine entsprechende Nahrungsbeschaffung hin. Neben den Beutegreifern, Aasfressern, Partikel- und Suspensionsfressern bilden die Filtrierer eine interessante Es gibt allerdings zahlreiche Beispiele, in denen diese Kombination nicht vorhanden ist und die dennoch Filtrierer gewesen zu sein scheinen. Eine Diskussion dieser Dinge würde hier aber zu weit führen. Es bleibt festzuhalten daß nach Fortey und Owens Trilobiten mit auffällig breiten, eventuell zusätzlich perforierten Cephalonsäumen in Verbindung mit konvexen Kopfschilden als Filtrierer anzusehen sind, die innerhalb einer Filtrierkammer, gebildet aus dem Kopfschild, eine "Nährlösung" verarbeiteten, die sie selbst aus der Wassersäule bzw. aus absichtlich aufgewirbeltem und dadurch im Wasser erneut gelöstem Sediment gebildet hatten. Die Perforationen am Kopfschild dienten, insoweit vorhanden, als Austritts- bzw. Eintrittsöffnungen für Wasser und nicht freßbares Sediment.
Ebenfalls anzumerken sind die Vertreter der Cheiruroidea, die zwar ein konterminantes Hypostom besitzen, das sich allerdings deutlich nach unten abneigt, in der Regel eine Granulation aufweist, keine Gabeln oder ähnliches besitzt, wie es für die Beutefresser angenommen wird, und dessen Entwicklung man nur als konservativ bezeichnen kann, will heißen es hat sich im Laufe der Jahrmillionen kaum verändert. Es scheint wahrscheinlich, daß diese Trilobiten eine spezialisierte Art der Nahrungsaufnahme entwickelt hatten - was diese aber war, bleibt bislang rätselhaft und kann hier nicht näher erläutert werden. In jedem Falle besetzten die Trilobiten verschiedene, uns vielleicht bis heute teilweise unbekannte Nischen und dies ist sicherlich eines ihrer zahlreichen Geheimnisse, die ihren Erfolg begründeten.
Letzte Aktualisierung: Donnerstag, 31.01.2019 14:30 Startseite - Einführung - Erdzeitalter - Fossilisation - Morphologie - Rekonstruktion - Sehapparat - Einrollung - Häutung - Bestachelung - Wachstum - Ernährung - Reproduktion - Pathologie - Ordnungen - Ursprünge - Aussterben - Fundorte - Kontinentaldrift - Präparation - Galerien - Gastsammlung - Fälschungen! - Literatur - Links - Evolution - Kreationismus - Glossar - Impressum - Über den Autor |
 Die Frage, was Trilobiten fraßen und welche Strategien sie dabei verfolgten, läßt sich nur indirekt angehen, da uns durch die Tatsache, daß es sich um eine - ohne direkte Nachkommen - ausgestorbene Klasse handelt, auch hier Grenzen gesetzt sind (siehe:
Die Frage, was Trilobiten fraßen und welche Strategien sie dabei verfolgten, läßt sich nur indirekt angehen, da uns durch die Tatsache, daß es sich um eine - ohne direkte Nachkommen - ausgestorbene Klasse handelt, auch hier Grenzen gesetzt sind (siehe: 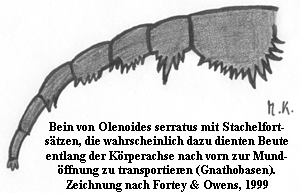 Trilobiten besaßen eine erstaunliche Diversität im Hinblick auf ihre äußere Form. Die morphologische Komplexität nahm vom Beginn des Kambriums an ins Ordovizium hinein beständig zu und bis ins Karbon war die Vielfalt noch bedeutend. Es ist nicht ungewöhnlich, innerhalb einer einzigen Biofazies auf bis zu 20 verschiedene Trilobitenspezies zu treffen, die sich morphologisch deutlich unterscheiden. Dies weist auf ein ganzes Spektrum von unterschiedlichen Nahrungsquellen hin, die es ermöglichten, daß diese Arten neben- und miteinander leben konnten, ohne sich gegenseitig die Nahrung streitig zu machen.
Trilobiten besaßen eine erstaunliche Diversität im Hinblick auf ihre äußere Form. Die morphologische Komplexität nahm vom Beginn des Kambriums an ins Ordovizium hinein beständig zu und bis ins Karbon war die Vielfalt noch bedeutend. Es ist nicht ungewöhnlich, innerhalb einer einzigen Biofazies auf bis zu 20 verschiedene Trilobitenspezies zu treffen, die sich morphologisch deutlich unterscheiden. Dies weist auf ein ganzes Spektrum von unterschiedlichen Nahrungsquellen hin, die es ermöglichten, daß diese Arten neben- und miteinander leben konnten, ohne sich gegenseitig die Nahrung streitig zu machen. Wir können und wollen nicht in alle Details der Ausführungen von Fortey und Owens einsteigen - insbesondere die parasitäre Option ist zu speziell - sondern uns auf für alle nachvollziehbare Grundmuster beziehen:
Wir können und wollen nicht in alle Details der Ausführungen von Fortey und Owens einsteigen - insbesondere die parasitäre Option ist zu speziell - sondern uns auf für alle nachvollziehbare Grundmuster beziehen: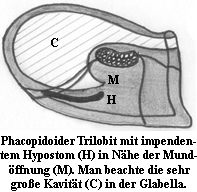 den größeren Vertretern der Klasse Trilobita gerechnet werden und eine recht konvexe Glabella besitzen. Diese und andere Beobachtungen führen Fortey und Owens zu der Annahme, daß es sich bei Trilobiten mit konterminantem oder impendentem Hypostom in der Regel um Beutegreifer oder Aasfresser handelte, die in der Lage waren mit Hilfe der
den größeren Vertretern der Klasse Trilobita gerechnet werden und eine recht konvexe Glabella besitzen. Diese und andere Beobachtungen führen Fortey und Owens zu der Annahme, daß es sich bei Trilobiten mit konterminantem oder impendentem Hypostom in der Regel um Beutegreifer oder Aasfresser handelte, die in der Lage waren mit Hilfe der  Im Gegensatz dazu hat man bei Trilobiten mit natanten, also nicht mit der Rostralplatte oder der Dublüre verbundenen Hypostomen eine auffällige Beständigkeit festgestellt, was die Ausformung desselben angeht. Diese hat sich in den gesamten 300 Millionen Jahren der Trilobitenherrschaft nur unwesentlich verändert. Diese Hypostome liegen gewissermaßen frei und sind wohl nur von mehr oder weniger weichem Gewebe umgeben gewesen, was sich nicht dazu geeignet hätte, sich wehrende Beutetiere zu fixieren oder kompakte Nahrung fachgerecht zu bearbeiten. Andererseits hat dies aber wohl auch eine gesteuerte Bewegung des Hypostoms begünstigt, in Art einer Klappe, die durch angesetzte Muskeln nach oben in Richtung der Mundöffnung fuhr und dabei half freßbare Partikel vom oder aus dem Sediment zuzuführen. Trilobiten, die dieser Art der Nahrungsaufnahme fröhnten, sind in der Regel kleinwüchsig und treten an ihren Fundorten bisweilen in großen Massen auf (bestes Beispiel:
Im Gegensatz dazu hat man bei Trilobiten mit natanten, also nicht mit der Rostralplatte oder der Dublüre verbundenen Hypostomen eine auffällige Beständigkeit festgestellt, was die Ausformung desselben angeht. Diese hat sich in den gesamten 300 Millionen Jahren der Trilobitenherrschaft nur unwesentlich verändert. Diese Hypostome liegen gewissermaßen frei und sind wohl nur von mehr oder weniger weichem Gewebe umgeben gewesen, was sich nicht dazu geeignet hätte, sich wehrende Beutetiere zu fixieren oder kompakte Nahrung fachgerecht zu bearbeiten. Andererseits hat dies aber wohl auch eine gesteuerte Bewegung des Hypostoms begünstigt, in Art einer Klappe, die durch angesetzte Muskeln nach oben in Richtung der Mundöffnung fuhr und dabei half freßbare Partikel vom oder aus dem Sediment zuzuführen. Trilobiten, die dieser Art der Nahrungsaufnahme fröhnten, sind in der Regel kleinwüchsig und treten an ihren Fundorten bisweilen in großen Massen auf (bestes Beispiel:  Gruppe. Trilobiten dieser Art hatten in der Regel einen großen Cephalonsaum der bisweilen regelrecht perforiert war, wie zum Beispiel bei den Vertretern der Gattungen
Gruppe. Trilobiten dieser Art hatten in der Regel einen großen Cephalonsaum der bisweilen regelrecht perforiert war, wie zum Beispiel bei den Vertretern der Gattungen  Problematisch stellen sich wieder einmal die Vertreter der
Problematisch stellen sich wieder einmal die Vertreter der